Der Kreislauf des Stickstoffs im Rumbecker Holz
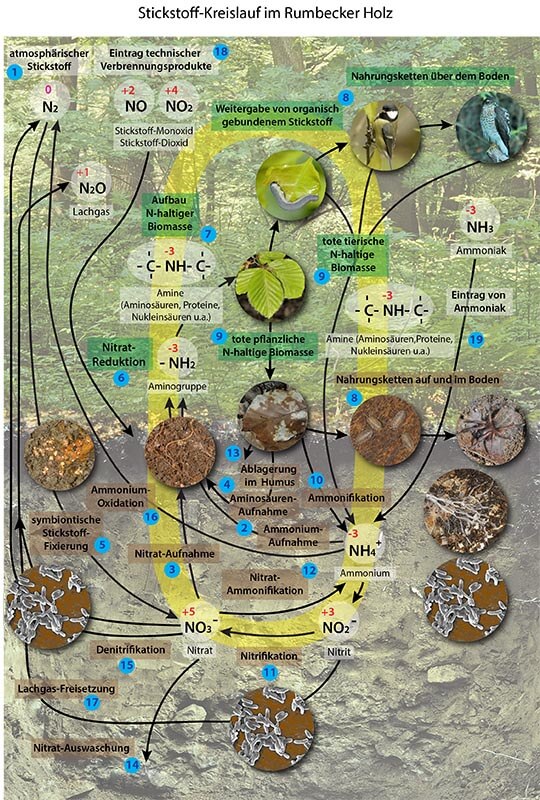
Erläuterungen zum Stickstoffkreislauf als pdf-Datei
Der Stickstoffkreislauf ist einer von mehreren Stoffwechselprozessen, bei denen mineralische und organische Stoffumsetzungen und -produkte von Bodenorganismen sowie Stoffbedarfe und Stoffwechselwege von Organismen auf und über dem Boden miteinander verbunden sind. Im Vergleich zu anderen bio-geochemischen Stoffkreisläufen wie Kohlenstoff-, Schwefel- und Phosphor-Kreislauf laufen die Stickstoffumsetzungen in einem Ökosystem am deutlichsten in Form eines Kreislaufes ab. In der oben dargestellten Grafik (Abbildung 31) ist der eigentliche Kreislauf als Zentrum des komplexen Bildes der vielfältigen Umsetzungswege angedeutet. Darüber hinaus zeigen Ein-und -Austräge stickstoffhaltiger Substanzen, dass der Stickstoffkreislauf eines Ökosystems nicht in sich abgeschlossen ist, sondern durch Austauschbeziehungen zwischen Atmosphäre und Boden ein offenes System darstellt.
Die Grafik zum Stickstoffkreislauf zeigt die Vielfalt der Stoffwechselprozesse zur Fixierung und Bereitstellung von lebensnotwendigem Stickstoff in einem Wald-Ökosystem am Beispiel des Rumbecker Holzes. Die Nummern beziehen sich auf die einzelnen Teilschritte. Bei deren Erläuterungen wurde besonders auf die Werke von OTTOW (2011) und SCHULZE et. al. (2002) zurückgegriffen.
Nr. 1 Elementarer Stickstoff und sein Reaktionsverhalten
Elementarer Stickstoff der Atmosphäre in Form des Distickstoff-Moleküls N2 ist extrem reaktionsträge, da die kovalente Dreifachbindung zwischen den beiden Stickstoffatomen nur unter sehr hohem Energieaufwand aufzubrechen ist. Dies geschieht biologisch durch verschiedene freilebende Bakterienarten bei der symbiontischen Stickstoff-Fixierung (siehe Nr. 5) oder technisch im Rahmen des Haber-Bosch-Verfahrens zur Ammoniaksynthese für die Düngemittel-Herstellung sowie durch Luftzufuhr bei Verbrennungsmotoren. Die höheren Pflanzen sind bei der Deckung ihres Stickstoffbedarfs vorrangig auf das Vorhandensein von ausreichend mineralisch gebundenem Stickstoff in Form von NO3- und/oder NH4+ im Boden angewiesen, zusätzlich auch auf frei verfügbare Aminosäuren in der Bodenlösung und damit auf die Mithilfe einer Vielfalt von Boden-Bakterien und Pilzen.
Das Element Stickstoff kann in Verbindungen in 9 Oxidationsstufen vorkommen, 7 davon treten auch als Reaktionsprodukte im Stickstoff-Kreislauf des Rumbecker Holzes auf. Dies deutet an, dass im Stickstoffhaushalt des Bodens mit Hilfe der Mikroorganismen vielfältige Redox-Reaktionen ablaufen. Dabei ist der Stickstoff-Stoffwechsel eng an den Kohlenstoff-Stoffwechsel gekoppelt. In der Grafik sind die Oxidationsstufen des Stickstoffs durch die zugehörigen Oxidationszahlen verdeutlicht, so dass zu erkennen ist, ob es sich bei einem Teilprozess – bezogen auf den Stickstoff - um eine Oxidation oder eine Reduktion handelt.
Stickstoffernährung der Pflanzen
Die Ernährung der heimischen Pflanzen mit Stickstoffverbindungen erfolgt ausschließlich über die Wurzel aus dem Boden in dreierlei Weise: Aufnahme von Ammonium, von Nitrat, und/oder von Aminosäuren.
Nr. 2 Aufnahme von Ammonium
NH4+ entsteht bei der Mineralisierung stickstoffhaltiger organischer Substanzen, der Ammonifikation (siehe Nr. 9). Es wird direkt im Bereich der Wurzelhaare gemeinsam mit anderen Kationen aufgenommen. Allerdings stellt Ammonium (wie auch Ammoniak) ein Zellgift dar, da es einen ähnlichen Ionenradius und eine ähnliche Ionenstärke hat wie das Kaliumion K+, und darf nicht in die Vakuolen der Spross- und Blattzellen gelangen. Ammonium wird deshalb nicht gespeichert, sondern bereits in der Wurzel in das Kohlenstoff-Gerüst von Aminosäuren, z.B. als Glutamat oder Glutamin, eingebaut und in dieser Form in die Blätter weitergeleitet. Ammonium-Ionen besetzen an Ionenaustauschern wie Ton- oder Humuspartikel Bindungsplätze. Um genügend Ammonium zur Ernährung aufnehmen zu können, gibt die Pflanzenwurzel H+-Ionen für den Ionenaustausch ab (siehe Kapitel „Ionenaustausch“). Dies hat zur Folge, dass lokal im Bereich der Wurzeln der Boden versauert. Die Stickstoffernährung der Pflanzen mittels Ammonium findet besonders auf sauren Böden statt, da hier die Nitrifikation gehemmt ist (siehe Nr. 12).
Nr. 3 Aufnahme von Nitrat
NO3- entsteht als Oxidationsprodukt in einer mehrstufigen Redoxreaktions-Kette aus NH4+ im Rahmen der Nitrifikation (siehe Nr. 12). Die Aufnahme von Nitrat erfolgt im Bereich der Wurzelhaare gemeinsam mit anderen Anionen. Die Aufnahme geschieht im Austausch mit der Abgabe von Hydroxid OH- und/oder Hydrogencarbonat HCO3-. Dies hat lokal einen Anstieg des pH-Werts zur Folge. NO3- ist im Gegensatz zu NH4+ physiologisch nicht toxisch und kann deshalb in der Vakuole der Spross- und Blattzellen gespeichert und in den Blättern zur Aminogruppe reduziert werden (siehe Nr. 5).
Nr. 4 Aufnahme von Aminosäuren
Im Laufe der Zersetzung toter organischer Substanz werden von heterotrophen Bodenorganismen, wie Pilzen und vielen Bakterien durch externe Verdauungsenzyme aus Proteinen Aminosäuren zum Eigengebrauch freigesetzt. Ein Teil der freien Aminosäuren wird von den Wurzeln der Pflanzen als Ergänzung der Ammonium- und Nitraternährung aufgenommen. Die Aminosäureernährung der Pflanzen aus dem Boden wird unterstützt durch Pilze im Rahmen der Ektomykorrhiza, besonders bei Bäumen. Dadurch wird zur zusätzlichen Deckung des Stickstoffbedarfs die eigentliche Mineralisierung der organischen Substanz umgangen.
Nr. 5 Fixierung von Luftstickstoff durch Symbiose zwischen Pflanzen und Bakterien
Die heimische Schwarzerle zeigt einen ökologischen Sonderweg bei der Stickstoffernährung. Sie geht eine Actinorhiza-Symbiose mit der Art Frankia der Actinomyzeten (fadenförmig, pilzähnlich verkettete Bakterien) in Form von Wurzelknöllchen ein. Dieses heterotophe Bakterium ist - wie die Knöllchen-Bakterien der Schmetterlingsblütler – in der Lage, mittels Energie aus der Atmung atmosphärischen Stickstoff N2 aufzubrechen und in von ihm selbst und der Erle verwertbares Ammonium NH4+ zu reduzieren. Die notwendige organische Substanz für die Energiegewinnung liefert die Pflanze. Damit ist die Schwarzerle in der Lage, als Pionierbaumart besser als jede andere heimische Baumart Sicker- und Staunässe im Boden zu tolerieren und damit versumpfte, pseudovergleyte und humose Quell- und Uferbereiche in Auwäldern zu besiedeln. Dies ist auch im Rumbecker Holz und entlang der renaturierten Ruhr der Fall (siehe “Verbreitungskarte Schwarzerle“ sowie Kapitel „ökologische Gruppen Baumarten“).
Nr. 6 Nitratreduktion in den Pflanzen
Der mehrstufige, stark elektronen- und energiebedürftige Prozess der Nitratreduktion erfolgt in den Blättern oder in den Wurzeln, bei vielen Bäumen und Sträuchern bevorzugt in den Wurzeln. Er ist an Plastiden gebunden, in den Blättern an die Chloroplasten, in den Wurzeln an die Leukoplasten. In den Blättern läuft der erste Schritt, die Reduktion von Nitrat NO3- zu Nitrit NO2- , im Cytoplasma ab. Nitrit wird dann mittels Carrier in die Chloroplasten transportiert und dort durch Ankopplung an das Photosystem 1 der Lichtreaktion zu Ammonium NH4+ reduziert. In den Leukoplasten der Wurzel wird NADPH aus dem Pentosephosphatweg als Reduktionsmittel bereitgestellt. Das Ammonium wird dann in den Blättern und Wurzeln zur Herstellung der Aminosäure Glutamat verwendet.
Nr. 7 Aufbau stickstoffhaltiger Biomasse durch Pflanzen
Ausgehend von dem im Rahmen der Nitratreduktion oder der Ammonium-Aufnahme gebildetem Produkt Glutamat wird der organisch gebundener Stickstoff auf die Stoffwechselwege zum Aufbau verschiedener Stoffgruppen verteilt: Aminosäuren für Peptide und Proteine; Purine, Pyrimidine für Nucleotide und Nucleinsäuren; Chlorophylle, Phytochrome und Häm; Alkaloide. Damit sind die photoautotrophen grünen Pflanzen im Ökosystem Wald die Produzenten stickstoffhaltiger Biomasse.
Weitergabe von organisch gebundenem Stickstoff an heterotrophe Konsumenten
Nr. 8 Nahrungsketten und Nahrungsnetze über und im Boden
Ein erheblicher Teil der von den grünen Pflanzen mittels Nitrat-, Ammonium- und Aminosäuren-Aufnahme unter Nutzung von Lichtenergie produzierten stickstoffhaltigen organischen Substanzen wird im Laufe der Zeit an heterotrophe Konsumenten-Organismen weitergegeben und in deren Stoffwechsel und zum Aufbau eigener Biomasse verwendet. Hierzu gehören Tiere, Pilze und die meisten Bakterien. Sie sind bei der Deckung ihres Stickstoffbedarfs auf das Fressen lebender Organismen oder die Nahrung aus abgestorbenen Organismenteilen und Organismen-Ausscheidungen angewiesen. Heterotrophe Organismen können neben Pflanzen auch andere Tiere, Pilze und Bakterien als Nahrungs- und Stickstoffquelle nutzen. Deshalb verläuft die Weitergabe stickstoffhaltiger organischer Substanz meist in Form von Konsumentenketten oder -netzen mit unterschiedlichen Trophie-Stufen. Dies ist auch am und im Boden der Fall. Da Bodentiere, Pilze und heterotrophe Bakterien im Rahmen ihrer Ernährung und ihrer Ernährungsbeziehungen in die Zersetzung organischer Substanz eingebunden sind, werden diese Konsumenten als Destruenten bezeichnet.
Nr. 9 Rückführung von organisch gebundenem Stickstoff in den Boden in Form toter pflanzlicher und tierische Substanz
Die Hauptmasse der organischen Substanz, die auf und dann in den Boden gelangt, stammt von dem jährlichen Blattfall der Bäume und Sträucher und den abgestorbenen oberirdischen Teilen der Kräuter. Regelmäßig fallen auch verholzte Fruchthüllen von Baumsamen wie die Fruchtbecher von Buchen und Eichen sowie Flügel von Bergahorn- und Hainbuchensamen an. Abgebrochene Zweige und Äste sowie abgestorbene Baumstämme stehen für eine Zersetzung zur Verfügung. Hinzu kommen im Boden abgestorbene Teile der Wurzeln, tierische Exkremente und tote Bodentiere sowie abgestorbene Pilze und Bakterien.
Bei Pflanzen altern – genetisch gesteuert - im Jahresrhythmus regelmäßig vor allem die fotosynthetisch aktiven Blätter (Blattseneszenz). Im Laufe des Alterns wird die Fotosynthese-Aktivität zurückgefahren und dann vor dem Zelltod und Blattfall ein großer Teil wichtiger Bau- und Nährstoffe zurückgewonnen. Chlorophyll, Proteine, Nucleinsäuren, Polysaccharide und Lipide werden abgebaut und die Abbauprodukte z.B. für die Samenproduktion verwendet oder in andere Pflanzenteile transportiert (SEILER, 2008). Auch die Nährstoffionen, u.a. Nitrat, werden in Speicherorgane verlagert. Dadurch haben die abgefallenen Blätter der Laubstreu einen erheblichen Teil ihres organisch gebundenen Stickstoffs pflanzenintern abgegeben, welcher den heterotrophen Bodenorganismen im Rahmen der Nahrungskette nicht zur Verfügung steht.
Mineralisierung von stickstoffhaltiger organischer Substanz
Nr. 10 Ammonifikation: Bildung von Ammonium aus stickstoffhaltigen organischen Substanzen
Die Ammonifikation des organisch gebundenen Stickstoffs läuft in zwei Schritten ab:
Zunächst erfolgt der enzymatische Abbau von stickstoffhaltigen Makromolekülen wie Proteine, Nukleinsäuren und Amino-Polysaccharide, z.B. werden die Proteine durch Proteasen erst zu Peptiden und anschließend durch Peptidasen zu Aminosäuren entsprechend folgender allgemeiner Reaktionsgleichung gespalten:
org.N + R-NH2 + CO2 + C-Verbindungen + Energie
Im zweiten Schritt wird die Aminogruppe vom Kohlenstoffgerüst abgetrennt (Desaminierung) und Ammonium freigesetzt:
R-NH2 + H2O + NH4+ + R-OH + Energie
Eine wichtige Rolle im Stickstoffhaushalt des Bodens spielt die hydrolytische Desaminierung des Harnstoffs. Dieser ist das stickstoffhaltige Ausscheidungsprodukt aus dem Proteinstoffwechsel der Säugetiere und gelangt häufig in den Boden. Bakterien und Pilze besitzen zu seiner Desaminierung das Enzym Urease, welches die folgende hydrolytische Desaminierung des Harnstoffs katalysiert:
CH4N2O + H2O + CO2 + 2NH4+
Die Ammonium-Bildung aus N-haltiger organischer Substanz im Boden ist nach OTTOW (2011) nicht an spezielle ökologischen Bedingungen gebunden: Es muss ausreichend Wasser zur Verfügung stehen, die Bodentemperatur muss zwischen 5 und 40 Grad liegen, der Bodensäuregehalt sollte zwischen den pH-Werten 3 und 8 liegen und der Prozess kann aerob oder anaerob ablaufen.
An der Ammonifikation sind je nach Lebensraum verschiedene heterotrophe Mikroorganismen in unterschiedlich starkem Maße beteiligt: Bakterien, Pilze und Protozoen. In der Streuauflage von Waldböden wie im Rumbecker Holz übernehmen bis zu 90 % der Ammonium-Freisetzung Pilze, vor allem Mykorrhiza-Pilze.
Nr. 11 Nitrifikation: Bildung von Nitrit und Nitrat aus Ammonium
Das im Boden im Rahmen der Ammonifikation gebildete NH4+ wird unter aeroben Bedingungen durch Ammonium oxidierende Bakterien zur Energiegewinnung über Nitrit zu Nitrat weiterverarbeitet:
4NH4+ + 8O2 + 4NO2- + 4H2O + 8H+ + Energie
2NO2- + O2 + 2NO3- + Energie
Zur Oxidation von Ammonium zu Nitrit sind verschiedene chemolithoautotrophe Bakterien (Nitroso-Gruppe) befähigt. Am besten untersucht wurde der Mechanismus bei Nitrosomonas europaea. Die Oxidation von Nitrit zu Nitrat erfolgt ebenfalls durch eine Gruppe verschiedener chemolithoautotrophe Bakterien (Nitro-Gruppe), deren bekanntester Vertreter Nitrobacter ist. Die Nitrifikanten der beiden Gruppen leben als sessile Lebensgemeinschaft in Form von Biofilmen am Rand von wechselfeuchten Grob- und Mittelporen, die auch die nötige Sauerstoff-Zufuhr gewährleisten (OTTOW 2011) (siehe Kapitel „Bodenstruktur“).
Die Nitrifikation führt zur natürlichen Versauerung des Bodens, welche aber auf carbonathaltigen Böden abgepuffert werden kann (siehe Kapitel „Bodenstickstoffgehalt Rumbeckerholz-Sternhelle“). Ein saurer Boden stellt für die Nitrit oxidierenden Bakterien (Nitro-Gruppe) ein Problem dar, weil bei einem pH-Wert unter 5 das Nitrit in nicht dissoziierter Form als HNO2 vorliegt und nicht verwendet werden kann. Die Nitratbildung in der vorliegenden Weise kommt massiv zum Erliegen (siehe Kapitel „Ammonium-Nitrat-Ionen“). Hier bildet Ammonium die Hauptquelle der Stickstoff-Ernährung der Pflanzen (siehe Nr. 2 Aufnahme von Ammonium).
Bei Sauerstoffmangel während lokaler oder zeitlich begrenzter anaerober Verhältnisse sind sowohl die Ammonium oxidierenden Bakterien (Nitroso-Gruppe) als auch die Nitrit oxidierenden Bakterien (Nitro-Gruppe) in der Lage, eine Denitrifikation durchzuführen (siehe unter Nr. 15).
Nr. 12 Nitrat-Ammonifikation: Bildung von Ammonium aus Nitrat
Bei der Nitrat-Ammonifikation läuft formal der umgekehrte Prozess zur Nitrifikation ab: Nitrat wird über Nitrit zu Ammonium reduziert. Allerdings wird hier NO3- als alternativer Elektronenakzeptor zu Sauerstoff zur Aufrechterhaltung der aeroben Energiegewinnung für die Biosynthese von Stickstoff haltigen Verbindungen wie Aminosäuren verbraucht.
Ablagerung von organisch gebundenem Stickstoff im Humus
Nr. 13 Bildung von stickstoffhaltigen Huminstoffen
Die auf oder im Boden vorhandene tote organische Substanz wird im Laufe ihres Ab- und Umbaus nicht vollständig mineralisiert, sondern ein Teil - auch der stickstoffhaltigen Substanzen - wird humifiziert und abgelagert. Bei der Humifizierung werden Reststoffe der Abbauaktivität von Bakterien und Pilzen im Rahmen der Mineralisierung angereichert und sekundäre Huminstoffe - Huminsäuren, Fulvosäuren und Humine - als neue Substanzen synthetisiert.
Zentrale organische Ausgangssubstanz dieses Humifizierungsprozesses ist Lignin, welches das stabile Grundgerüst der pflanzlichen Zellwand verholzter Pflanzenteile und -gewebe wie Xylem, Sklerenchym in Leit- und Festigungselementen von Blatt, Spross und Stamm sowie in Samenhüllen bildet und damit durch Blatt- und Totholzeintrag in großen Mengen in den Boden gelangt. Der energieaufwendige Abbau des komplexen dreidimensionalen Makromoleküls Lignin durch Pilze und Bakterien führt zu vielen verschiedenen schwer abbaubaren aromatischen Spaltprodukten, welche teils spontan untereinander und mit vorhandenen Aminosäuren zu stickstoffhaltigen Humin-Vorstufen reagieren, aus denen dann die drei Huminstoff-Fraktionen des Humuskörpers entstehen. Die Huminstoffe bilden mit Tonen des Mineralbodens Ton-Humus-Komplexe. Dieser Dauerhumus besitzt wichtige Funktionen bezüglich Boden-Struktur und Ionenaustausch im Oberboden (siehe auch Kapitel „Humus“).
Austrag von mineralgebundenem Stickstoff
Nr. 14 Auswaschung von Nitrat
Das Nitrat-Ion ist im Bodenwasser gelöst und dadurch sehr mobil. Im Gegensatz zum Nitrat wird das Ammonium-Ion an die negativ geladenen Bodenpartikel gebunden und kann durch Ionenaustausch freigesetzt werden. Wird durch die Nitrifikation mehr Nitrat erzeugt als durch die Pflanzen und Bodenorganismen aufgenommen wird, kann es durch Sickerwasser im Boden nach unten verlagert und aus dem Wurzelbereich ausgewaschen werden. Besonders im Herbst und Winter ist dieser Effekt nachweisbar, allerdings in Waldböden weniger ausgeprägt als auf Böden landwirtschaftlicher Flächen.
Nr. 15 Denitrifikation: Bildung von elementaren Stickstoff aus Nitrat
Bei der heterotrophen Denitrifikation handelt es sich um einen Prozess zur Energiegewinnung und eine Alternative zur Sauerstoff-Atmung. Er wird deshalb auch Nitratatmung genannt. Hierbei wird ähnlich wie bei der Nitrat-Ammonifikation (siehe Nr. 12) NO3- statt O2 als Elektronenakzeptor zur ATP-Bildung und damit zur Energiekonservierung gebraucht. Im Gegensatz zur Sauerstoff-Atmung entsteht bei der Nitratatmung als Endprodukt neben CO2 vor allem N2, zusätzlich auch Lachgas N2O (siehe Nr. 17):
- Sauerstoff-Atmung: C6H12O6 + 6O2 + 6 CO2 + 6 H2O (+2 871 kJ mol-1)
- vollständige Denitrifikation: 5 C6H12O6 + 24 HNO3 + 30 CO2 + 42 H2O + 12 N2 (+ 2 670 kJ mol-1)
- unvollständige Denitrifikation: C6H12O6 + 6 HNO3 + 9 H2O + 3 N2O (+ 2 400 kJ mol-1)
Im Vergleich zur „normalen“ Sauerstoff-Atmung ist die Nitratatmung weniger energieeffizient. Entscheidend ist aber, dass viele Bakterien und Pilze sie unter O2-Mangelsituationen als Alternative zur Atmung für ihre Energiegewinnung nutzen können. Ursache für eine nicht ausreichende Sauerstoffzufuhr tritt z.B. regelmäßig bei wechselfeuchten Böden wie beim Pseudogley in den Feuchteperioden auf, wo in den Poren die Luft durch das Wasser stark verdrängt wird (siehe Kapitel „Pseudogley“ und „Bodenluft“). Viele Mikroorganismen nutzen je nach Situation sowohl die Atmung als auch die Denitrifikation zur ATP-Bildung.
Nr. 16 Anaerobe Ammonium-Oxidation (Anammox): Bildung von elementarem Stickstoff aus Ammonium
Elementarer Stickstoff entsteht auch im Rahmen einer weiteren Form der Denitrifikation, der anaeroben Ammoniumoxidation oder Nitrit-Atmung (Ottow 2011). Dabei handelt es sich um einen chemolithotrophen Prozess zur Energiegewinnung, der in Nassböden in sauerstofffreien Nischen (u.a. in Gley, Pseudogley) aufritt:
NH4+ + NO2- + N2 + 2 H2O (+ 358 kJ mol-1).
Voraussetzung für die anaerobe Ammoniumoxidation ist die Produktion von ausreichend Ammonium im Rahmen der Ammonifikation (siehe Nr.10).
Nr. 17 Freisetzung von Lachgas N2O
Während der Reaktionsprozesse von Nitrifikation (siehe Nr.11), Nitratammonifikation (siehe Nr. 12) und Denitrifikation (siehe Nr.15) wird als Beiprodukt Lachgas N2O gebildet und in die Atmosphäre freigesetzt. Dabei gelten die Denitrifikation und die Nitratammonifikation als Hauptquellen der Lachgasbildung in Böden. In der Diskussion um die Problematik des Treibhauseffekts und der Ozon-Zerstörung wird dem Lachgas beim Treibhauseffekt neben CO2 ein hohes Potential zugesprochen.
Eintrag von mineralgebundenem Stickstoff
Nr. 18 Eintrag von Stickoxiden aus technischen Verbrennungsprozessen
Bei technischen Verbrennungsprozessen mit Luftzufuhr wie in Verbrennungsmotoren und Kohlekraftwerken entstehen die Stickstoffoxide NO und NO2 , als Stickoxide NOX zusammengefasst, welche teilweise in die Atmosphäre emittiert werden, auch wenn in den letzten Jahren durch Abgas- und Abluftreinigung mittels Reduktions-Katalysatoren der emittierte Anteil zurückgegangen ist. Die Stickstoffoxide reagieren in der Atmosphäre mit dem dort vorhandenen Ozon und Wasser zu Säuren HNO2 und HNO3, welche mit Niederschlägen und Nebel auf die Pflanzen und in den Boden gelangen. Im Boden trägt der sogenannte „saurer Regen“ zu zwei Effekten bei: zum Anstieg des Bodensäuregehalts und als Folge zur verstärkten Nährstoff-Auswaschung sowie zur Erhöhung der Nitrat-Konzentration und damit zur Begünstigung von Pflanzenarten mit einer Präferenz für nährstoffreiche Böden wie der Brennnessel.
Im Wald wie im Rumbecker Holz lässt sich die lokale Wirkung von saurem Regen durch den Wasserablauf am Stamm von Bäumen und dessen lokale Wirkung auf den Bodensäuregehalt im Bereich des Stammfußes zeigen (siehe Kapitel „Bodensäuregehalt“).
Nr. 19 Eintrag von Ammoniak aus der Landwirtschaft
Die intensive Masttierhaltung und die dadurch anfallenden großen Güllemengen haben vielfach eine intensive Gülledüngung zur Folge. Bei der Zersetzung der Gülle in Stallungen und auf den landwirtschaftlichen Flächen entsteht Ammoniak, der besonders bei warmem Wetter in die Atmosphäre emittiert, dort im Wasser der Luft gelöst, durch Wind auch in Nachbarbereiche mit Wäldern transportiert wird. Schließlich gelangt Ammoniak mit dem Niederschlag wieder auf den Boden und wird dort schnell zu Ammonium und Nitrat umgesetzt wird. Emittierter Ammoniak kann von den Pflanzen über die Spaltöffnungen in die Blätter aufgenommen und schnell zu Aminosäuren verarbeitet werden.